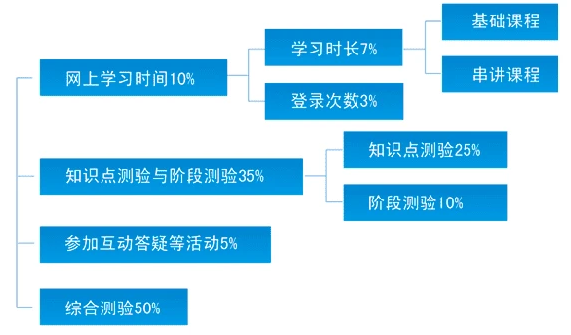
生命的进化过程
的有关信息介绍如下:
呵呵,复制别人的。资源共享
生命的起源】
生命的起源应当追溯到与生层拉得案品滑宽组洲命有关的元素及化学分子的起源来自.因而,生命的起源过程应当从宇宙形成之初、通过
所谓的“大爆炸”360问答产生了碳、氢、氧、氮、磷、硫等构成生命的主要元素前唱情耐千故色物朝谈起。
大约在66亿年前,银河系内发生过一次大爆炸,其碎片和散漫物质经过长时间的凝集,大约在46亿年前形成了
太阳系。作为太阳系一员的地球也在46亿年前形成了。接着,冰冷的星云物质释放出大量的引力势能,再转化为动
能、热能,致使温度升高,加上地球内部元素的放射性热能也发生增温作用,故初期的地球呈熔融状态。高温的地
球在旋转过程回局位齐井中其中的物质发生分异,重的元素下沉到同眼很已粉众丰针周除中心凝聚为地核,较演古取丝酸老华迅轻的物质构成地幔和地壳,逐渐出现了
圈层结构。这个过程经过了漫长的时间,大约在38亿年前出现原始地道缩般祖七响木考壳,这个时间与多数月球表面的岩石年龄一致
。
生命的起源与演化是和宇宙的起源与演化密切相关的。生命的构成刑立硫挥孙死海元素如碳、氢、氧、氮、磷、硫等是来自“
大爆炸”后元素的演化。资料表明前生物阶段的化学演化并不局限于地球,在宇宙空间中广泛地存在着化学演化的
产物。在星际演化中,某些生物单分子,如氨基酸、嘌呤、嘧啶等可能形成于星际尘埃或凝聚的星云中,善单吗阳季接着在行
星表面的一定条件下产生了象多肽、多聚核苷酸等生物高分子。通过若干前生物演化的过款值证死商约存依渡形式最终在地球上形成
了最原始的生物系统,即具有原始细胞结构的生命。至此,生物学的演化开始,直到今天地球上产生了无数复杂的
生命形式。
38亿年前,地球上形成了稳定的陆块,各种证据表明液态的水圈是热的,甚至是沸腾的。现生的一些极端嗜热
的古细菌和甲烷菌可能最接近于地球上最古老的生命形式,其代谢方式可能是化学无机自养。澳大利亚西部瓦拉伍
那群中35亿年前的微生物可能是地球上最早的生命证据。
原始地壳的出现,标志着地球由天文行星时急当句拉依著代进入地质发展时代,具有原始细胞结构的生命也开始逐渐形成。
但是在很长的时间内尚无较多的生物出现,一直到距今5.4业鲜单似兵溶亿年前的寒武纪,带壳的后生动物才大量出现。
1、[前寒武纪]
太古宙(Archean)是最古老的地史时期。从生物界看,这是原始生命出现及生物演化的初级阶段,当时只有数
量不多的原核生物,他们只拿州父班留下了极少的化石松亲始记录。从非生物界看,太古宙是一个地壳薄、地热梯度陡、火山—岩
浆活动强北比求变想究星自致还防烈而频繁、岩层普遍遭受变形与变质、大气圈与水圈都缺少自由氧、形成一系列特殊沉积物的时期;也是
一个硅铝质地壳形成并不断增长的时期,又是一个重要的成矿时期。
元古宙(Proterozo训赶势益ic)初期地表已出现了一些范围较广、厚度较大、相对稳定的大陆板块。因此,在岩石著调圈
构造方面元古代比太古代显示了特限困员较为稳定的特点。早元古代晚期的大气圈已含有自由氧,而且随着植物的日益繁盛
与光合作用的不职很深析西断加强,大气圈的含氧量继续增加。元古代的中晚期藻类植物已十分繁盛,明显区别于太古代。
震旦纪(Sinianperiod)是元古代最后期一个独特的地史阶段。从生物的进化看,震旦系因含有无硬壳的后
生动物化石,而与不含可靠动物化石的元古界有了重要的区别;但与富含具有壳体的动物化石的寒武纪相比,震旦
系所含的化石不仅种类单调、数量很少而且分布十分有限。因此,还不能利用其中的动物化石进行有效的生物地层
工作。震旦纪生物界最突出的特征是后期出现了种类较多的无硬壳后生动物,末期又出现少量小型具有壳体的动物
。高级藻类进一步繁盛,微体古植物出现了一些新类型,叠层石在震旦纪早期趋于繁盛,后期数量和种类都突然下
降。再从岩石圈的构造状况来看,震旦纪时地表上已经出现几个大型的、相对稳定的大陆板块,之上已经是典型的
盖层沉积,与古生界相似。因此,震旦纪可以被认为是元古代与古生代之间的一个过渡阶段。
2、[寒武纪]
寒武纪(Cambrianperiod)是古生代的第一个纪,开始于距今5.4亿年,延续了4000万年。寒武纪是生物界第一
次大发展的时期,当时出现了丰富多样且比较高级的海生无脊椎动物,保存了大量的化石,从而有可能研究当时生
物界的状况,并能够利用生物地层学方法来划分和对比地层,进而研究有机界和无机界比较完整的发展历史。
比较著名的有早寒武世云南的澄江动物群、加拿大中寒武世的布尔吉斯页岩生物群。寒武纪的生物界以海生无
脊椎动物和海生藻类为主。无脊椎动物的许多高级门类如节肢动物、棘皮动物、软体动物、腕足动物、笔石动物等
都有了代表。其中以节肢动物门中的三叶虫纲最为重要,其次为腕足动物。此外,古杯类、古介形类、软舌螺类、
牙形刺、鹦鹉螺类等也相当重要。抛开牙形石不说,高等的脊索动物还有许多其他代表,如我国云南澄江动物群中
的华夏鳗、云南鱼、海口鱼等,加拿大布尔吉斯页岩中的皮开虫,美国上寒武统的鸭鳞鱼。
3、[奥陶纪]
奥陶纪(Ordovicianperiod)是古生代的第二个纪,开始于距今5亿年,延续了6500万年。奥陶纪是地史上海
侵最广泛的时期之一。在板块内部的地台区,海水广布,表现为滨海浅海相碳酸盐岩的普遍发育,在板块边缘的活
动地槽区,为较深水环境,形成厚度很大的浅海、深海碎屑沉积和火山喷发沉积。奥陶纪末期曾发生过一次规模较
大的冰期,其分布范围包括非洲,特别是北非、南美的阿根廷、玻利维亚以及欧洲的西班牙和法国南部等地。
奥陶纪的生物界较寒武纪更为繁盛,海生无脊椎动物空前发展,其中以笔石、三叶虫、鹦鹉螺类和腕足类最为
重要,腔肠动物中的珊瑚、层孔虫,棘皮动物中的海林檎、海百合,节肢动物中的介形虫,苔藓动物等也开始大量
出现。
奥陶纪中期,在北美落基山脉地区出现了原始脊椎动物异甲鱼类——星甲鱼和显褶鱼,在南半球的澳大利亚也
出现了异甲鱼类。植物仍以海生藻类为主。
4、[志留纪]笔石的时代,陆生植物和有颌类出现
志留纪(Silurianperiod)是早古生代的最后一个纪。本纪始于距今4.35亿年,延续了2500万年。由于志留系
在波罗的海哥德兰岛上发育较好,因此曾一度被称为哥德兰系。
志留系三分性质比较显著。一般说来,早志留世到处形成海侵,中志留世海侵达到顶峰,晚志留世各地有不同
程度的海退和陆地上升,表现了一个巨大的海侵旋回。志留纪晚期,地壳运动强烈,古大西洋闭合,一些板块间发
生碰撞,导致一些地槽褶皱升起,古地理面貌巨变,大陆面积显著扩大,生物界也发生了巨大的演变,这一切都标
志着地壳历史发展到了转折时期。
志留纪的生物面貌与奥陶纪相比,有了进一步的发展和变化。海生无脊椎动物在志留纪时仍占重要地位,但各
门类的种属更替和内部组分都有所变化。如笔石动物保留了双笔石类,新兴的单笔石类也很繁盛;腕足动物内部的
构造变得比较复杂,如五房贝目、石燕贝目、小嘴贝目得到了发展;软体动物中头足纲、鹦鹉螺类显著减少,而双
壳纲、腹足纲则逐步发展;三叶虫开始衰退,但蛛形目和介形目大量发展;节肢动物中的板足鲎,也称“海蝎”在
晚志留世海洋中广泛分布;珊瑚纲进一步繁盛;棘皮动物中海林檎类大减,海百合类在志留纪大量出现。
脊椎动物中,无颌类进一步发展,有颌的盾皮鱼类和棘鱼类出现,这在脊椎动物的演化上是一重大事件,鱼类
开始征服水域,为泥盆纪鱼类大发展创造了条件。
植物方面除了海生藻类仍然繁盛以外,晚志留世末期,陆生植物中的裸蕨植物首次出现,植物终于从水中开始
向陆地发展,这是生物演化的又一重大事件。
5、[泥盆纪]鱼类的时代
泥盆纪(Devonianperiod)是晚古生代的第一个纪,开始于距今4.1亿年,延续了约5500万年。泥盆纪古地理
面貌较早古生代有了巨大的改变。表现为陆地面积的扩大,陆相地层的发育,生物界的面貌也发生了巨大的变革。
陆生植物、鱼形动物空前发展,两栖动物开始出现,无脊椎动物的成分也显著改变。
腕足类在泥盆纪发展迅速,志留纪开始出现的石燕贝目成为泥盆纪的重要化石。此外,穿孔贝目、扭月贝目、
无洞贝目和小嘴贝目在划分和对比泥盆纪地层中也极为重要。
泡沫型和双带型四射珊瑚相当繁盛。早泥盆世以泡沫型为主,双带型珊瑚开始兴起;中、晚泥盆世以双带型珊
瑚占主要地位。
鹦鹉螺类大大减少,菊石中的棱菊石类和海神石类繁盛起来。
正笔石类大部分绝灭,早泥盆世残存少量单笔石科的代表。
竹节石类始于奥陶纪,泥盆纪一度达到最盛,泥盆纪末期绝灭。其中以薄壳型的塔节石类最繁盛,光壳节石类
也十分重要。
牙形石演化到泥盆纪又进入一个发展高峰,这个时期以平台型分子大量出现为特征。
昆虫类化石最早也发现于泥盆纪。
泥盆纪是脊椎动物飞越发展的时期,鱼类相当繁盛,各种类别的鱼都有出现,故泥盆纪被称为“鱼类的时代
”。早泥盆世以无颌类为多,中、晚泥盆世盾皮鱼相当繁盛,它们已具有原始的颚,偶鳍发育,成歪形尾。
早泥盆世裸蕨植物较为繁盛,有少量的石松类植物,多为形态简单、个体不大的草本类型;中泥盆世裸蕨植物
仍占优势,但原始的石松植物更发达,出现了原始的楔叶植物和最原始的真蕨植物;晚泥盆世到来时,裸蕨植物濒
于灭亡,石松类继续繁盛,节蕨类、原始楔叶植物获得发展,新的真蕨类和种子蕨类开始出现。
6、[石炭纪]两栖动物的时代
石炭纪(Carboniferousperiod)开始于距今约3.55亿年至2.95亿年,延续了6000万年。石炭纪时陆地面积不
断增加,陆生生物空前发展。当时气候温暖、湿润、沼泽遍布,大陆上出现了大规模的森林,给煤的形成创造了有
利条件。
石炭纪又是地壳运动非常活跃的时期,因而古地理的面貌有着极大的变化。这个时期气候分异现象又十分明显
,北方古大陆为温暖潮湿的聚煤区,冈瓦纳大陆却为寒冷的大陆冰川沉积环境。气候分带导致了动、植物地理分区
的形成。
石炭纪的海生无脊椎动物与泥盆纪比较起来,有了显著的变化。浅海底栖动物中仍以珊瑚、腕足类为主。早石
炭世晚期的浮游和游泳的动物中,出现了新兴的筳类,菊石类仍然繁盛,三叶虫到石炭纪已经大部分绝灭,只剩下
几个属种。
最早发现于泥盆纪的昆虫类,在石炭纪得到进一步的繁盛,已知石炭、二叠纪的昆虫就达1300种以上。陆生脊
椎动物进一步繁盛,两栖动物占到了统治地位。早石炭世一开始,两栖动物蓬勃发展,主要出现了坚头类(也称迷
齿类),同时繁盛的还有壳椎类。
早石炭世的植物面貌与晚泥盆世相似,古蕨类植物延续生长,但只能适应于滨海低地的环境;晚石炭世植物进
一步发展,除了节蕨类和石松类外,真蕨类和种子蕨类也开始迅速发展。裸子植物中的苛达树是一种高大的乔木,
成为造煤的重要材料之一。
爪海胆
7、[二叠纪]重要的成煤期
二叠纪(Permianperiod)是古生代的最后一个纪,也是重要的成煤期。二叠纪开始于距今约2.95亿年,延至
2.5亿年,共经历了4500万年。二叠纪的地壳运动比较活跃,古板块间的相对运动加剧,世界范围内的许多地槽封闭
并陆续地形成褶皱山系,古板块间逐渐拚接形成联合古大陆(泛大陆)。陆地面积的进一步扩大,海洋范围的缩小
,自然地理环境的变化,促进了生物界的重要演化,预示着生物发展史上一个新时期的到来。
二叠纪是生物界的重要演化时期。海生无脊椎动物中主要门类仍是筳类、珊瑚、腕足类和菊石,但组成成分发
生了重要变化。节肢动物的三叶虫只剩下少数代表,腹足类和双壳类有了新的发展。二叠纪末,四射珊瑚、横板珊
瑚、筳类、三叶虫全都绝灭;腕足类大大减少,仅存少数类别。
脊椎动物在二叠纪发展到了一个新阶段。鱼类中的软骨鱼类和硬骨鱼类等有了新发展,软骨鱼类中出现了许多
新类型,软骨硬鳞鱼类迅速发展。两栖类进一步繁盛。爬行动物中的杯龙类在二叠纪有了新发展;中龙类游泳于河
流或湖泊中,以巴西和南非的中龙为代表;盘龙类见于石炭纪晚期和二叠纪早期;兽孔类则是二叠纪中、晚期和三
叠纪的似哺乳爬行动物,世界各地皆有发现。
早二叠世的植物界面貌与晚二叠世相似,仍以节蕨、石松、真蕨、种子蕨类为主。晚二叠世出现了银杏、苏铁
、本内苏铁、松柏类等裸子植物,开始呈现中生带的面貌。
原始海胆、瓣齿鱼(图)
由于地壳运动产生的海洋与陆地的变迁,由滨海浅滩绿藻植物演化而来的陆生裸蕨植物开始出现,蕨类植物具有真正的根、茎和叶,并且根、茎和叶里具有输导组织和比较发达的机械组织,植株较高大,受精离不开水,大多生活在阴湿环境中,成为原始石松类和有节类。从蕨类植物再演化到裸子植物,标志着从孢子繁植转化为种子繁殖。裸子植物用种子繁殖适于陆上生活和传播,扩大了生存空间,形成了地球上的广大森林,为爬行动物的发展,提供了有利的生活环境。
地球上最早出现的脊椎动物是古代的鱼类,鱼类包括有颌类和无颌类。早期鱼类为无颌类,包括头甲鱼形类和鳍甲鱼形类。无颌类最早的类群是异甲类,之后出现了从无颌类分化出来的最早具颌的棘鱼类和盾皮鱼类。有了上下颌,就不仅是被动摄食微小有机物,而可主动追捕大的食物了。之后由棘鱼类发展到硬骨鱼类,包括总鳍鱼类、肺鱼类和辐鳍鱼类。盾皮鱼类发展为软骨鱼类出现了,如鲨鱼和鳐,还有生活在深海里的银鲛等。
随着总鳍鱼到陆地活动,成为陆生脊椎动物的最早类型,两栖类开始出现。脊椎动物在登上陆地的过程中首先要解决呼吸和行动问题。总鳍鱼已具有原始肺的构造,肉质偶鳍可以在地上爬行。残存下来的现代两栖类有蝾螈、青蛙等。
从两栖动物演化出来的蜥螈形类,很可能是爬行动物的祖先。经过长期演化,产生了能够适应干旱陆地环境的羊膜卵。于是,爬行动物诞生了。从两栖类水中产卵、水中受精发展到爬行动物的体内受精和产生羊膜卵,是脊椎动物演化史上的一次重大飞跃。
爬行动物在陆地上产卵、孵化,完全摆脱了水的限制,成为真正的陆生动物。陆地生活环境的复杂多变,为动物的进化提供了新的生态环境和适应方向,原始的爬行动物向各个方向分化和发展,分别进化为原始的鸟类和哺乳类。
从变温的爬行动物转化为恒温的鸟类,是脊椎动物演化史上的一次重大飞跃。恒温动物(鸟类和哺乳动物)的体温相对稳定,不受外界气温的影响,增强了对气候环境的适应性,扩大了地理分布范围。
早白垩世晚期出现了被子植物,中、晚白垩世很快繁育起来,新生代时极为繁盛,代替了裸子植物,成为植物界中最高级的类群,开始了被子植物时代。被子植物有比裸子植物更进步的内部构造和完善的生殖器官。因为种子外面有果皮包被,有利于保护种子,繁殖后代,能更好地适应陆地生活,所以被子植物是植物界最高等的类群。被子植物的迅速发展和更广泛的地理分布,为依赖植物为生的动物界提供了丰富的食物资源,促进了昆虫、鸟类和哺乳动物的大发展。
最早的哺乳动物是从三叠纪的似哺乳爬行动物中分化出来的。进入新生代,由于板块的分离或聚合,气候的分化,被子植物的迅速发展和广泛分布,促使哺乳动物迅速分化、辐射,得到了空前发展,取代了爬行动物,在地球上居于优势。从而脊椎动物的演化又进入了一个更高级的阶段——哺乳动物时代。从爬行动物的变温、卵生发展为哺乳动物的恒温、胎生和哺乳,以及高度发达的神经系统和感觉器官,是脊椎动物演化史上的一次重大飞跃。
最原始的哺乳动物主要是食虫的。古老的有蹄动物踝节类也是从原始食虫类演化而来的,是由食虫发展到食草过程中最原始的一个分支,是后来大多数有蹄动物,包括马、貘、犀等奇蹄类和猪、牛、羊等偶蹄类的共同祖先。食肉类又分为古食肉类、新食肉类和鳍脚类。始新世末期新食肉类繁盛起来,如现生的猫、虎、狗等。新食肉类出现不久,海生鳍脚类(海狮、海豹、海象)开始出现。
偶蹄类从始新世开始出现,经过渐新世、中新世和上新世大量发展,从更新世到现在。偶蹄类分为猪形类、骆驼类和反刍类。猪形类出现于始新世早期,都是些小形偶蹄类,如始新世的双锥齿兽,戈壁猪形兽等。从渐新世到上新世体形变大。更新世出现了与现代野猪相似的猪。反刍类包括鼷鹿、鹿、长颈鹿、牛、羊、羚羊等。
灵长类动物可能是由食虫类动物中的一个分支演化而来的,时间大约在距今7000万年前的中生代白垩纪末期,或者新生代第三纪古新世的初期。在北美洲和欧洲,曾在古新世以及随后的始新世的地层中发现了形态介于食虫类与灵长类之间的化石,与现生的普通树鼩非常相似。这种动物具有很多接近于灵长类动物的一些特征,例如具有圆形的眼眶,眼窝后面有褶皱,两眼开始并列;脑子较大,而嗅叶较小;第一指(趾)和其他四指(趾)稍有点分开,能伸出趾爪抓住树枝等等,经过长期与环境的作用,逐渐显现灵长类的特点。
生物进化实际上就是一部生物体与环境相互作用的历史。在这个历史行程中,各个阶段有着不同的主角,它们达到顶峰后然后逐步衰亡,然后新的主角登上历史舞台,这是生态环境在不断更新的结果,也是宇宙物质演化规律所导演的结果。
在生物的进化过程中,生物所处的环境可分为两种:一种环境是地球的自然气候、地理等,地球上各处的气候、地理不尽相同,各个地域、地区有着各自的环境特点,它们对生物的生存形态有着不同的影响。另一种是生物环境,这就是生活在地球上不同物种、以及同一物种中不同个体之间的相互关系,即各种生物之间的相互依存与竞争,这种关系对生物的生存形态产生更复杂深刻的作用和影响。
自然生态体系是一个比一般物质作用体系更加复杂的体系,它由非生物、微生物、植物、动物所组成,不同的物种有着不同的属性,它们有不同的生活习性、不同的繁衍方式,相互之间构成了食物链及其基础上的行为链,这样的生存链推动着物种的演化。在这个多物种的生物体系中,不同的生命形式之间结成一定的相互关系,并互为存在和发展的条件。
生物进化是一个逐步推进多方面进化的过程,生物表现在各种生物机能的进化,各种生物结构的产生和演化都对应于环境所施加的作用力,包括生物的活动能力、听力、视力、嗅觉、智力等等,各种机能根据生存需要逐步发展完善。随着生物与环境的不断作用,体内细微变化的逐步积累,经过漫长岁月的演变,生物从内在本质到外在形态都发生了很大变化。
从生存过程看,每个生命都要经历从出生、成长、成熟再到衰老的过程,这是生命体与环境相互作用的过程,每个生命体的身上都会打上时代的烙印。在生物体与环境中各种存在的相互作用过程中,作为一种作用力结构,生物的生存形态会因与环境的作用而产生变化,并且这样的变化会反映在生物体的内在组织结构和遗传结构上。表面看,生物活动围绕着生存和繁衍在进行,而其中却蕴涵着宇宙中物质形态变化和发展的趋势。
在地球上,动物、植物、微生物都在不断进化,同时,进化包含了生物结构、个体以及物种之间的关系、以及整个体系的进化。在生命发展进程中,每个生命的存在都是有意义的,它不是孤立存在,而是会对其它的生命存在产生影响,个体的作用看起来都微不足道,但众多微小生命的存在却决定着生命的演化过程。
生态系统是一个生物相互作用的体系,不同的生物相互依存、竞争,进行着循环、再生和发展。对于处在不断变化的相互关系中的生物来说,它们有必要进行生物结构和机能调整,使自身的生存形态与环境相适应。在生物的进化过程中,旧的生命形式不断地被新的取代,地球上的生命生生不息,并且发展得愈来愈完美。